








A prophage-encoded anti-phage defense system that prevents phage DNA packaging by targeting the terminase complex
Posted on: 19 May 2025 , updated on: 20 May 2025
Preprint posted on 3 March 2025
Defense mechanism inception! Listeria prophages protect their host bacteria with anti-phage systems, leaving them susceptible to their own defense systems…read how this may actually be an evolutionary advantage!
Selected by Briana Guzman, Noah Franz, UofA IMB565Categories: microbiology
Background:
Bacteriophages are viruses, categorized as either lytic or temperate, that infect bacterial cells and hijack their machinery for replication and propagation across cells. Lytic phages immediately package, replicate, and release their progeny via cell lysis while temperate phages persist in the bacterial chromosome and replicate along with the host in a lysogenic state1. In the presence of stressors, temperate phages are excised from the genome to escape the unfavorable environment known as prophage induction. In homeostatic conditions, lysogeny is thought to be a long-term physiological state where phage and bacteria share mutual interests and co-exist in some cases2.
Phages and bacteria are in a constant evolutionary arms race. Phages acquired unique strategies to avoid host defense mechanisms and propagate when their bacterial host may no longer provide the resources needed for survival. On the other hand, host bacteria evolved strategies to defend against invading predatory phages and may induce cell death to protect the larger population from phage spread. While these two entities are traditionally thought of as opposing forces in a constant predator-prey relationship, this study highlights an interesting example of phage-bacteria commensalism in Listeria monocytogenes.
Listeria monocytogenes (Lm) is an intracellular bacterial pathogen of mammals that can rearrange the plasma membrane to be engulfed by the host. Lm resides within vacuoles or phagosomes, depending on the cell type it has invaded, and is translocated to the cytosol for replication 3. The ability of Lm strains to house prophages in the genome is not well studied, specifically the overall impact of these “native” phages on bacterial virulence.
The authors of the preprint highlighted here previously revealed that Lm strain 10403S and its prophage, ϕ10403S have a protective effect on the bacteria during mammalian infection Transcriptomics data of ϕ10403S helped them identify a gene, orf65, that is constitutively transcribed under both phage states, lysogenic and lytic conditions. In this preprinted study, the authors sought to look further into the gene as it is one of few phages transcribed during lysogeny and only located within the early lytic modules. Specifically, they investigated the gene’s role in ϕ10403S switch between states.
Key Findings:
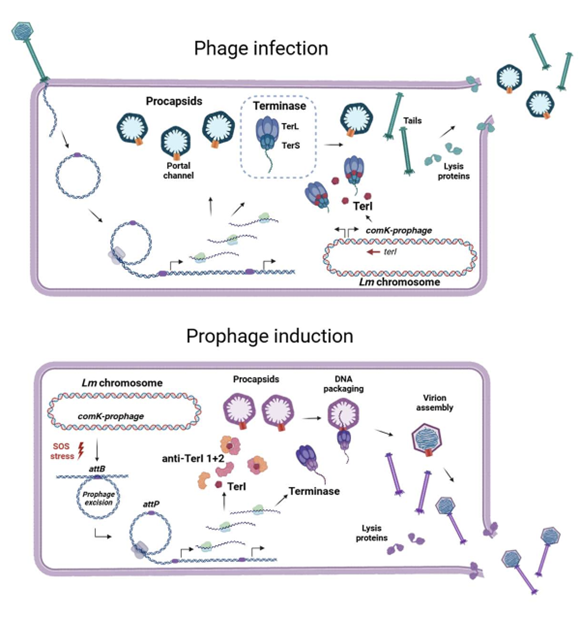
The Listeria monocytogenes (Lm) strain 10403S has a sole prophage ϕ10403S that supports the survival of Lm during mammalian cell infection. Transcriptome analysis of ϕ10403S indicated a gene that is constitutively transcribed under lysogenic and lytic conditions. This gene,orf65, is highly conserved among different Lm prophages. During lysogenic conditions (where orf65 remains dormant in the host genome), this gene does not appear to affect bacterial growth or virion production indicating it does not play a role in regulating lysogeny. However, when cells are stressed with mitomycin C, which induces lytic replication, overexpression of orf65 appeared to significantly reduce the number of virions produced. These findings indicate that orf65 likely plays some role in interfering with the lytic cycle of the prophage.
To understand the mechanism by which orf65 disrupts virion production, the authors used a Lm orf65 knockout strain and assessed the effects of knocking out this gene on phage DNA replication, gene transcription and cell lysis. Using a series of separate assays, the authors found that orf65 did not inhibit any of these functions. This paradoxical result led the researchers to look at processes further downstream, since they did already know that orf65 overexpression reduced the number of virions under lytic conditions. To address this point, they looked at the assembly of packaged virions using transmission electron microscopy (TEM). Upon scanning different bacterial sections, they found that wild-type bacteria exhibited heavily stained capsids, whereas bacteria overexpressing orf65 exhibited white “ghost-like” capsids. This led them to speculate that orf65 plays a role in packaging DNA into assembled phage capsids; an hypothesis which was later supported by using a DNase I protection assay in which only encapsulated DNA (properly packaged) was protected from degradation.
These findings raise an interesting question about the role of orf65: why would a prophage express a gene that directly disrupts its own DNA packaging? The authors speculate that this intrinsic feature allows the prophage to protect the host bacteria from other invading phages during lysogeny. However, if this were true, the prophage must have some counter-defense system to allow its own productive replication and assembly of virions during its lytic life-cycle. In search for such a system, the authors screened for mutations in ϕ10403S that rescue virion production upon overexpression of orf65. Sequencing the genome of 17 independent lysogens recovered from this screen, they found them all to carry the same mutation, a single nucleotide substitution in the operator site of Cro. Considering Cro is required for the expression of early genes in the ϕ10403S genome, the authors independently screened these early genes and found two genes, LMRG_01518 and LMRG_02984 which could rescue virion production in the presence of orf65 overexpression.
Lastly, having discovered an anti-phage (orf65) as well as a counter-defense system (LMRG_01518 and LMRG_02984), the authors investigated how widespread these systems are among other Lm prophages. They found the promoter region of orf65 to be present in 96% of the 149 Lm prophages they examined. Furthermore, the authors found a protein that associates with LMRG_01518 and plays a role in the uncovered counter-defense strategy which is present in 30.9% of the strains they analyzed. These findings reveal that targeting DNA packaging as an anti-phage defense system is a common strategy of Lm prophages. Moreover, this strategy has likely evolved multiple times independently to generate unique counter-defense systems.
Model from paper:
Why we chose to highlight this preprint:
- This study reveals a novel, symbiotic strategy used by Lm prophages as a counter defense system to evade exogenous phages without compromising the lytic lifestyle of the prophage.
- It provides insight into a unique system that exemplifies how a phage-bacterial relationship can function for the protection of both.
- Shows a new system that supports the idea of prophages strategically aligning their genomes to develop defense systems within.
Questions for the authors:
- At the end of the first figure, you show that ORF65’s ability to block virion production was not specific to 1040S, but to temperate phages since lytic phages seemed to have no virion production effect by orf65-expression. Did you look at any similar genomic sequences in lytic phages and compare it to the orf65 gene locus? For example, did you look at a specific attachment site (attP) in a lytic phage genome? Maybe the differences in effects of virion production could be further explained by comparing the genome sequences.
- The amount of phage DNA recovered from capsids produced by orf65-expressing bacteria was found to be significantly lower than that recovered from capsids produced by bacteria not expressing orf65. Do you think partial expression of or65 in bacteria would result in partial reduction of capsids recovered? Could there be a certain limit of orf65 needed to reach this phenotype?
- No DNA was detected in procapsids produced by the ΔterS mutant compared to the capsids produced by orf65-expressing bacteria which had less capsid production than bacteria not expressing orf65. How do you explain that both ΔterSL mutant and bacteria expressing orf65 showed small capsids of 40nm that did not present internal staining, but have what appears to be significantly different capsid production (fig. 2F)?
- Bacteria were infected with a ϕ10403S-phage that carries a kanamycin resistance gene to allow for selection of lysogens upon phage induction specifically triggered by UV radiation. Why did you choose to switch to using UV radiation as a stress condition instead of Mitomycin C like in previous experiments? Do you think that if you compared different bacterial growth media components known to induce phage production, like lithium chloride or nalidixic acid, you would see similar results?
The wider context of this study:
The prophage in this study encodes an anti-phage defense system which inhibits its own success as a lytic virion. While this may seem paradoxical, this mechanism appears to be evolutionarily conserved with 96% of analyzed Listeria prophage sequences containing a promoter sequence for anti-phage defense systems. Considering this, it appears that limiting the prophage’s own success as a lytic virion may lead to an evolutionary advantage. This advantage may be explained by the prophage protecting the host, and subsequently itself from other invading lytic phage, a tradeoff strategy that unintentionally acts in favor of both the prophage and Listeria. Furthermore, endogenous prophages may acquire specialized “self counter-defense” systems to enhance their lytic potential and avoid their own encoded anti-phage system. However, these strategies seem to be variable among prophage populations and not as well conserved, questioning their necessity for prophage success.
These insights bring an interesting new perspective to phage biology and phage-host interactions, expanding our traditional understanding of “host immunity” and highlighting conserved mechanisms for prophage survival. Further understanding the machinery used by phages to persist within their host could also give insight for developing therapeutic approaches using phage therapy to treat bacterial infections and prevent bacterial spread.
References:
- Feiner, R., Argov, T., Rabinovich, L. et al. A new perspective on lysogeny: prophages as active regulatory switches of bacteria. Nat Rev Microbiol 13, 641–650 (2015)
- Stern, Adi, and Rotem Sorek. “The phage-host arms race: shaping the evolution of microbes.” BioEssays : news and reviews in molecular, cellular and developmental biology vol. 33,1 (2011): 43-51.
- Pizarro-Cerdá, Javier, and Pascale Cossart. “Listeria Monocytogenes: Cell Biology of Invasion and Intracellular Growth.” Microbiology Spectrum, vol. 6, no. 6 (2018).
doi: https://doi.org/10.1242/prelights.40498
Read preprint (No Ratings Yet)
(No Ratings Yet)Sign up to customise the site to your preferences and to receive alerts
Register hereAlso in the microbiology category:
Circadian Clock Programming of Anticipatory Antiviral Immunity Gates Enteric Virus Infection Susceptibility
Owen Ang
Inhibition of VP2-mediated entry: a potential antiviral strategy to treat or prevent calicivirus disease
Orestis Savva
Gut microbiome changes over the course of multiple sclerosis differentially influence autoimmune neuroinflammation
Carole Djagang et al.
preLists in the microbiology category:
SciELO preprints – From 2025 onwards
SciELO has become a cornerstone of open, multilingual scholarly communication across Latin America. Its preprint server, SciELO preprints, is expanding the global reach of preprinted research from the region (for more information, see our interview with Carolina Tanigushi). This preList brings together biological, English language SciELO preprints to help readers discover emerging work from the Global South. By highlighting these preprints in one place, we aim to support visibility, encourage early feedback, and showcase the vibrant research communities contributing to SciELO’s open science ecosystem.
| List by | Carolina Tanigushi |
BioMalPar XVI: Biology and Pathology of the Malaria Parasite
[under construction] Preprints presented at the (fully virtual) EMBL BioMalPar XVI, 17-18 May 2020 #emblmalaria
| List by | Dey Lab, Samantha Seah |
1
ECFG15 – Fungal biology
Preprints presented at 15th European Conference on Fungal Genetics 17-20 February 2020 Rome
| List by | Hiral Shah |
EMBL Seeing is Believing – Imaging the Molecular Processes of Life
Preprints discussed at the 2019 edition of Seeing is Believing, at EMBL Heidelberg from the 9th-12th October 2019
| List by | Dey Lab |
Antimicrobials: Discovery, clinical use, and development of resistance
Preprints that describe the discovery of new antimicrobials and any improvements made regarding their clinical use. Includes preprints that detail the factors affecting antimicrobial selection and the development of antimicrobial resistance.
| List by | Zhang-He Goh |