








Unlocking the secrets of kangaroo locomotor energetics: Postural adaptations underpin increased tendon stress in hopping kangaroos
Posted on: 25 March 2024
Preprint posted on 8 February 2024
Article now published in eLife at http://dx.doi.org/10.7554/eLife.96437.3
Should we all be hopping like a kangaroo? @LivEvoBiomech , @SamRRCross and @james_charles90 review this exciting preprint: Unlocking the secrets of kangaroo locomotor energetics.
Selected by EMB EMB_Liv, James Charles, Roger KissaneCategories: physiology, zoology
PreLight Authors
Samuel Cross, James Charles, Roger Kissane
Introduction
Terrestrial mammals generally experience higher metabolic rates the faster they run (1). Large to moderately sized macropods (kangaroos, wallabies) are known to decouple this relationship between locomotor speed and cost, as they are uniquely capable of hopping at greater speeds without incurring significantly higher energy expenditure (2; Figure 1). This ability is related to their capacity to generate greater stress in the Achilles tendon with speed, which in turn allows for improved storage of elastic strain energy and a greater contribution of tendinous recoil to total work performed at the ankle as they hop faster (3). However, while this phenomenon is well documented, its underlying mechanistic causes remain unclear.
Thornton et al., hypothesize that postural modulation (i.e. changing posture with speed) may provide a mechanism by which tendon stress can be increased. Unlike most mammals of comparable size, large macropods move with highly flexed lower limb postures where the ankle operates at a relatively low mechanical advantage (EMA) (Figure 2). A low MA should increase the stress on the tendon, allowing for greater storage of elastic strain energy. Therefore, they theorise that proportional reductions to MA with increasing speed may allow for increased tendon stress and elastic storage, which would facilitate the decoupling of energetic expenditure and movement velocity.
In this study, the authors generate 3D musculoskeletal models of kangaroos (Figure 2) and integrate 3D kinematics and force plate data from freely hopping kangaroos. These musculoskeletal models allow for quantitative analysis of kangaroo posture (Figure 2a) and EMA (Figure 2b) which are evaluated across differing speeds.
 Figure 1: Energetic expenditure versus speed in terrestrial mammals. Most mammals (blue) show an increase in energy consumption with increasing speed, whereas the relationship is flat in macropods (red). Data is hypothetical/illustrative. (A) running mammals, from (1); (B) Red kangaroos, from (2).
Figure 1: Energetic expenditure versus speed in terrestrial mammals. Most mammals (blue) show an increase in energy consumption with increasing speed, whereas the relationship is flat in macropods (red). Data is hypothetical/illustrative. (A) running mammals, from (1); (B) Red kangaroos, from (2).
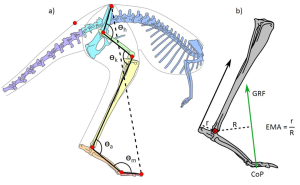
Figure 2: Illustration of the kangaroo model (a) and the calculation of effective mechanical advantage (EMA) (b).
Key Findings
Kangaroo posture changes with speed
Thornton et al., showed that kangaroos change posture with increasing speed and body mass. However, while all limb joints became increasingly flexed with greater mass, increasing speed was associated only with localised flexion of the distal joints. This change in posture was found to decrease the leverage of the calves, causing MA to decrease with speed. Meanwhile, the forces within the ankle (the joint moment) increased with speed. These factors in combination led to relatively higher Achilles tendon stresses as kangaroos hopped faster.
Postural change increases elastic savings
Evaluation of work, or force generated over a certain distance, performed at the ankle found that while positive work (i.e. force produced during muscle contractions), increases with speed (similar to all other animals), kangaroos are also able to increase the negative work (i.e. force produced during muscle lengthening) performed. Typically, positive work is produced at the ankle during the push-off part of stance where the joints extend via muscle contractions, while the while negative work is generated at the start of stance, corresponding to the period where elastic energy is stored in the Achilles tendon as it lengthens. This increase in negative work with speed is an artefact of changing ankle posture, and allows for increased storage of elastic strain energy, which can be returned during the positive work phase replacing active (and metabolically costlier) muscle contractions. Because positive and negative work increased at equal rates with speed, the net work done about the ankle remained fairly constant irrespective of increasing speed. Thus, the authors conclude that postural modulation allows macropods to decouple locomotor cost and speed by improving elastic storage and utilisation (Figure 3).
Evolutionary implications for hopping
When compared to other mammals, the mechanical advantages used by kangaroos are disproportionately low for their body size. This means that even fairly marginal changes in mechanical advantage can lead to substantial increases in tendon stress and elastic storage. It illustrates a mechanistic difference between macropods and quadrupedal mammals, in which equivalent postural changes would incur only minimal increases in stress. However, such high tendon stresses present scaling challenges for macropods, as hopping at high speeds at large body sizes may dangerously approach or exceed the safety limit of the Achilles tendon. This suggests that the largest fossil kangaroos may have been incapable of gaits characteristic of their smaller-bodied contemporaries.
Our Opinions
This article attempts to address a longstanding (and in our opinion, very interesting) question in evolutionary biomechanics/physiology. The ability of large macropods to defy what is otherwise a fairly universal relationship between speed and cost in running mammals has been discussed at length in the technical literature, but (to our knowledge) no succinct theory regarding underlying mechanical causes has thus far been demonstrated. We therefore appreciate the integrative approach taken by the authors, combining experimental data with musculoskeletal modelling, as well as incorporating a large body of analyses. In addition, there has also been a great deal of discussion about the energetic penalties of flexed postures in terrestrial vertebrates (a topic several of us are keenly interested in), so it was enjoyable to read about a reversal to this rule.
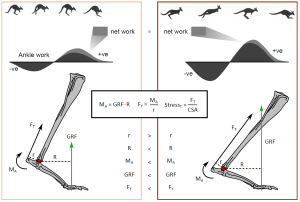
Figure 3: Summary figure from Thornton et al., illustrating how changes to posture and mechanical advantage can increase tendon stresses and elastic energy storage.
Questions for authors
- Throughout the study, the authors have used a mixture of macropodid taxa, and we wondered whether this may carry any further implications? For example, are there phylogenetic distinctions between red and grey kangaroos in any of the locomotor parameters reported here?
- How were the speeds allocated to the binning of Slow, Medium and Fast? Similarly with body mass, how were individuals classed as Small, Medium and Large?
- Are there any structural differences in the macropod Achilles tendon compared to other equally large mammals?
- Are there any further plans for the musculoskeletal models built for this study? Maybe some predictive simulations to further investigate the relationships between the Achilles tendon and locomotor cost, if muscle force-generating properties are available?
References
- Kram R. and Taylor, C. R. 1990. Energetics of running: a new perspective. Nature, 346, 265-267.
- Dawson, T. J. and Taylor, C. R. 1973. Energetic Cost of Locomotion in Kangaroos. Nature, 346, 313-314.
- Biewener, A. A., Konieczynski, D. D. and Baudinette, R. V. 1998. In Vivo Muscle Force–Length Behaviour During Steady-Speed Hopping in Tammar Wallabies. Journal of Experimental Biology, 201, 1681-1694.
doi: https://doi.org/10.1242/prelights.36914
Read preprint (No Ratings Yet)
(No Ratings Yet)Sign up to customise the site to your preferences and to receive alerts
Register hereAlso in the physiology category:
Inhibition of the gut ceramidase Asah2 decelerates the vertebrate ageing rate
Jeny Jose
Feeding and reproduction of a tropical coastal copepod across warming and copper gradients
Tina Nguyen
Resilience to cardiac aging in Greenland shark Somniosus microcephalus
Theodora Stougiannou
Also in the zoology category:
Behavioral characteristics of an extremely old rhesus macaque in a zoo: Dementia-like symptoms and implications for quality of life of geriatric animals
Stefan Friedrich Wirth
Heatwave winners and losers: cryptic coral holobionts differ in thermal tolerance
Maitri Manjunath
Consciously detecting and recognizing a past visual word after its sensory trace is gone
Maitri Manjunath
preLists in the physiology category:
Developmental regulation: molecular and ecological niches
This conference was held at the Station Biologique de Roscoff (France) and brought together researchers exploring how diverse niche environments shape developmental processes across scales. Spanning topics from ecological and metabolic influences to signalling networks, mechanics and gene regulation, the meeting highlighted the interplay between intrinsic and extrinsic factors in controlling cell fate and tissue organisation. This preList gathers preprints discussed by speakers and poster presenters during the meeting. Please do get in touch at preLights@biologists.com if you notice any relevant preprints that we may have missed.
| List by | Ingrid Tsang |
preLighters’ choice – Handpicked DevBio preprints
preLighters with expertise across developmental and stem cell biology have nominated a few developmental biology (and related) preprints they’re excited about and explain in a few paragraph why. Concise preprint highlights, prepared by the preLighter community – a quick way to spot upcoming trends, new methods and fresh ideas.
| List by | Theodora Stougiannou et al. |
Keystone Symposium on Stem Cell Models in Embryology 2026
The Keystone Symposium on Stem Cell Models in Embryology, 2026, was organised by Jun Wu (UT Southwestern), Jianping Fu (University of Michigan) and Miki Ebisuya (TU Dresden) and held at Asilomar Conference Grounds in California (US). The meeting discussed recent advances made in establishing stem-cell-based embryo models, what fundamental insights into developmental processes have been gleaned from them, as well as how they are beginning to be applied more widely. This prelist contains preprints by presenters at the talk and poster sessions at the conference, which our Reviews Editor in attendance spotted. Please do reach out to preLights@biologists.com if you notice any that we’ve missed.
| List by | Ingrid Tsang |
October in preprints – DevBio & Stem cell biology
Each month, preLighters with expertise across developmental and stem cell biology nominate a few recent developmental and stem cell biology (and related) preprints they’re excited about and explain in a single paragraph why. Short, snappy picks from working scientists — a quick way to spot fresh ideas, bold methods and papers worth reading in full. These preprints can all be found in the October preprint list published on the Node.
| List by | Deevitha Balasubramanian et al. |
Biologists @ 100 conference preList
This preList aims to capture all preprints being discussed at the Biologists @100 conference in Liverpool, UK, either as part of the poster sessions or the (flash/short/full-length) talks.
| List by | Reinier Prosee, Jonathan Townson |
Fibroblasts
The advances in fibroblast biology preList explores the recent discoveries and preprints of the fibroblast world. Get ready to immerse yourself with this list created for fibroblasts aficionados and lovers, and beyond. Here, my goal is to include preprints of fibroblast biology, heterogeneity, fate, extracellular matrix, behavior, topography, single-cell atlases, spatial transcriptomics, and their matrix!
| List by | Osvaldo Contreras |
FENS 2020
A collection of preprints presented during the virtual meeting of the Federation of European Neuroscience Societies (FENS) in 2020
| List by | Ana Dorrego-Rivas |
TAGC 2020
Preprints recently presented at the virtual Allied Genetics Conference, April 22-26, 2020. #TAGC20
| List by | Maiko Kitaoka et al. |
Autophagy
Preprints on autophagy and lysosomal degradation and its role in neurodegeneration and disease. Includes molecular mechanisms, upstream signalling and regulation as well as studies on pharmaceutical interventions to upregulate the process.
| List by | Sandra Malmgren Hill |
Cellular metabolism
A curated list of preprints related to cellular metabolism at Biorxiv by Pablo Ranea Robles from the Prelights community. Special interest on lipid metabolism, peroxisomes and mitochondria.
| List by | Pablo Ranea Robles |
Also in the zoology category:
SciELO preprints – From 2025 onwards
SciELO has become a cornerstone of open, multilingual scholarly communication across Latin America. Its preprint server, SciELO preprints, is expanding the global reach of preprinted research from the region (for more information, see our interview with Carolina Tanigushi). This preList brings together biological, English language SciELO preprints to help readers discover emerging work from the Global South. By highlighting these preprints in one place, we aim to support visibility, encourage early feedback, and showcase the vibrant research communities contributing to SciELO’s open science ecosystem.
| List by | Carolina Tanigushi |
Biologists @ 100 conference preList
This preList aims to capture all preprints being discussed at the Biologists @100 conference in Liverpool, UK, either as part of the poster sessions or the (flash/short/full-length) talks.
| List by | Reinier Prosee, Jonathan Townson |
Bats
A list of preprints dealing with the ecology, evolution and behavior of bats
| List by | Baheerathan Murugavel |