








Hibernating ribosomes tether to mitochondria as an adaptive response to cellular stress during glucose depletion
Posted on: 15 November 2023 , updated on: 27 November 2023
Preprint posted on 8 October 2023
Are ribosomes guarding mitochondria during starvation? The fascinating cryo-EM/ET images of ribosomes attached to mitochondria provided in this preprint suggest that they might be!
Selected by Barbora KnotkovaCategories: molecular biology
Background
During nutrient starvation, cells have limited energy to spare and therefore they shut down most anabolic processes. Only proteins whose function will help the cells overcome starvation will be translated under such conditions [1].This leaves the cell with many inactive ribosomes, which are targeted for degradation [2, 3]. However, a recent study examining septin assemblies in the fission yeast Schizosaccharomyces pombe (S. pombe) noted that cytosolic ribosomes accumulate on mitochondria in glucose-starved cells, but the authors did not explore this curious finding further [4]. Gemin and colleagues have now made this phenomenon the main subject of their research and present their findings in this preprint. They illuminate the molecular details underlying the sequestration of inactive ribosomes to mitochondria using cryo-electron microscopy (EM) methods in conjunction with biochemical tools.
Key findings
- Ribosomes enter a self-inhibition mode upon prolonged glucose depletion
-
- After 7 days of glucose depletion of S. pombe, no active ribosomes could be observed by polysome profiling. In addition, no mRNA or tRNA was found to be associated with purified ribosomes – as analysed by single particle cryoEM – suggesting that indeed no protein synthesis was taking place.
- The cryoEM structures of ribosomes from glucose-deprived yeast revealed that the P-site, where the peptide-bound tRNA usually interacts with the ribosome during protein synthesis, is blocked by a helix of the large ribosomal subunit. Furthermore, this conformational shift in the helix disrupts an interaction important for translation initiation.
- Prolonged glucose depletion induces ribosomal tethering to the outer mitochondrial membrane (see Figure below)
-
- Using cryo-electron tomography (cryoET), the research team could analyse the cellular structure of glucose-starved S. pombe. They saw that the mitochondrial network was fragmented, and that the resulting round mitochondria were richly decorated with ribosomes, as has been described before [4].
- To show that ribosomal tethering to the mitochondria is a consequence of glucose-starvation and not mitochondrial fragmentation, the authors inhibited mitochondrial fragmentation in the yeast cells by deleting the mitochondrial fission factor Dnm1. Just like in wild-type cells, ribosomes in Ddnm1 cells associated with the outer mitochondrial membrane upon prolonged glucose depletion, despite the mitochondria being elongated.
- Ribosomes are arranged into oligomers on the outer mitochondrial membrane and face the membrane with the small ribosomal subunit (see Figure below)
-
- The authors analysed the orientation of the ribosomes on the mitochondrial membrane by mapping sub-tomogram averaged electron densities of ribosomes back to the electron tomograms of cell slices.
- The ribosomes were attached to the membrane via the small ribosomal subunit, an orientation distinct from active ribosomes previously found on mitochondria [5, 6].
- The ribosomes formed organised clusters on the mitochondrial membrane with the help of previously unknown binding sites mediating dimer, trimer, tetramer and pentamer formation between ribosomes.
- Ribosomes interact with the outer mitochondrial membrane via the Cpc2/RACK1 subunit
-
- Sub-tomogram averaging of the cryo-tomograms could not provide molecular details about the connection between ribosomes and the outer mitochondrial membrane, probably due to high flexibility. The researchers therefore fitted the single-particle cryoEM structures of purified ribosomes into the tomograms.
- The fit placed the ribosome-associated protein Cpc2 in close proximity to the membrane. Indeed, a Dcpc2 deletion strain did not accumulate ribosomes on mitochondria upon prolonged glucose starvation, suggesting that Cpc2 is the mitochondria-interacting factor of ribosomes.
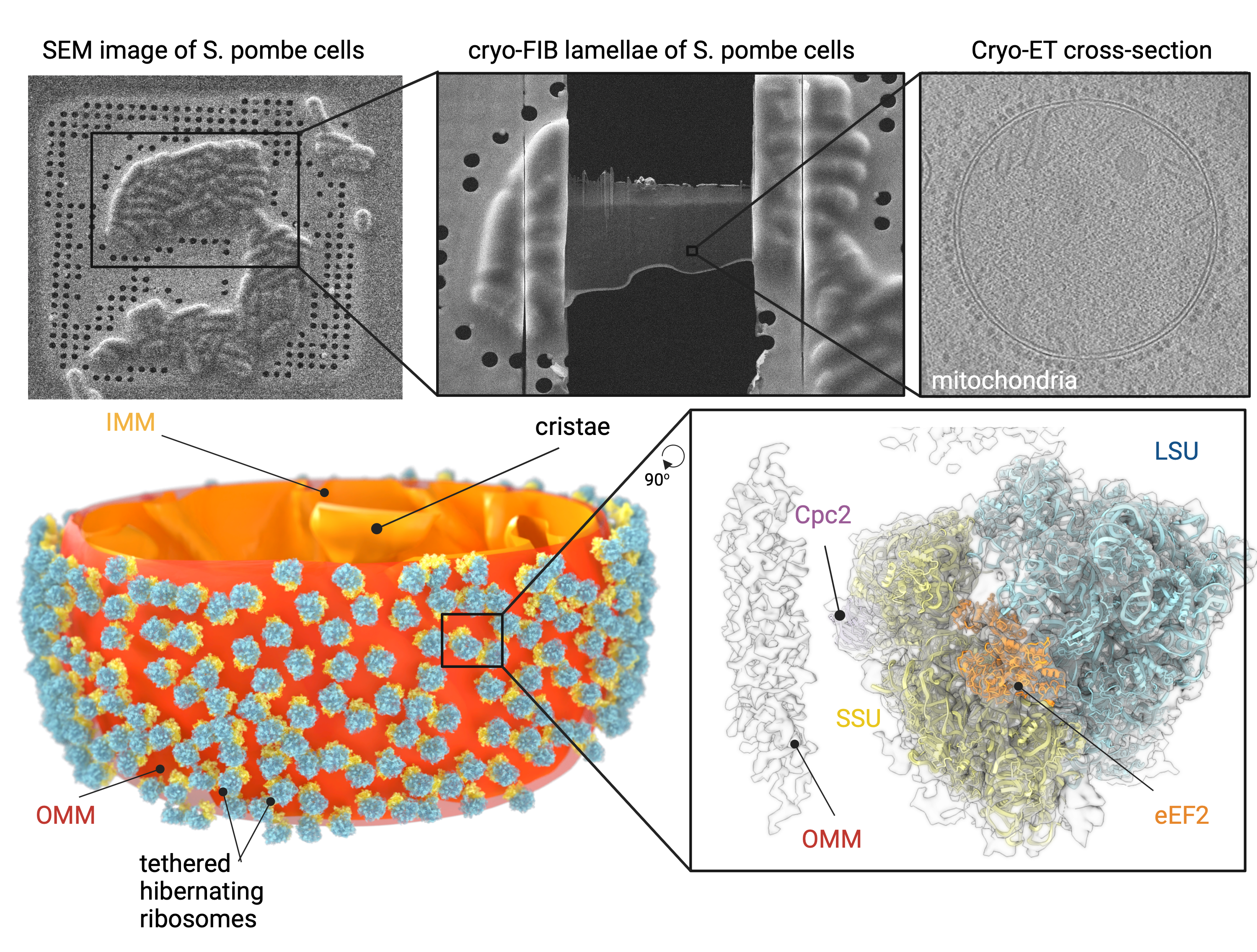
This figure shows how glucose-starved S. pombe cells were sectioned into thin slices by cryo-FIB (A), and how cryoET imaging of these revealed ribosome-decorated mitochondria. Through subtomogram averaging, the mitochondria-tethered ribosomes could be further resolved to determine the location of individual subunits within the ribosome (C). When these higher-resolution ribosomes were mapped back onto a 3D reconstruction of the mitochondrion, all of the ribosomes faced the mitochondrial membrane via their small subunit (coloured in yellow) (D). Figure adapted from the preprint.
Hypotheses for the function of hibernating ribosomes on mitochondria during glucose starvation:
- Ribosomes tethered to fragmented mitochondria protect them from mitophagy and stabilize the mitochondrial potential, thereby averting cell death.
- Mitochondria-bound ribosomes are waiting for the starvation to be over, at which time they can start to synthesise mitochondrial proteins, directly feeding them into mitochondria, which then will have plenty of resources to power the rest of the cell.
What I like about this preprint:
I really enjoyed reading this preprint because it examines such a fascinating phenomenon. I had never heard of ribosomes sitting on mitochondria before and found the cryoET images showing this rather impressive. I also liked that the preprint was not too long and was kept to the point. I hope that there will be more research on this topic and that we can find out more about what these ribosomes are doing on the mitochondria!
Questions and comments
- Do you have any ideas as to what Cpc2’s binding partner in the outer mitochondrial membrane might be? Would it be possible to map translocation complexes of the outer membrane onto the tomograms of mitochondria to determine whether the ribosomes are in close proximity to these, supporting your hypothesis that the hibernating ribosomes are ready to restart mitochondrial protein synthesis upon nutrient repletion?
- Mammalian mitochondria tend to fuse, rather than fragment, during starvation to protect themselves from mitophagy. Do you have an explanation for why pombe mitochondria do the opposite? Do other yeast species’ mitochondria behave the same as S. pombe’s during starvation?
- In your preprint, especially in the abstract, it sounds like you propose that the sequestration of ribosomes to mitochondria may confer cell survival. However, in the corresponding data, Cpc2 deletion impacted cell survival even in EMM 2% glucose media (Figure 4B), where cells are not glucose-deprived, mitochondria are not fragmented and ribosomes are not sequestered to mitochondria (Figure S9). Therefore Cpc2’s interaction with the mitochondrial membrane and the associated ribosome sequestration may not be the factors that confer cell survival, but rather it may be another function of Cpc2 important specifically in the EMM medium but not under glucose-deprivation.
References
1. Janapala, Y., T. Preiss, and N.E. Shirokikh, Control of Translation at the Initiation Phase During Glucose Starvation in Yeast. Int J Mol Sci, 2019. 20(16).
2. Kraft, C., et al., Mature ribosomes are selectively degraded upon starvation by an autophagy pathway requiring the Ubp3p/Bre5p ubiquitin protease. Nat Cell Biol, 2008. 10(5): p. 602-10.
3. An, H. and J.W. Harper, Systematic analysis of ribophagy in human cells reveals bystander flux during selective autophagy. Nat Cell Biol, 2018. 20(2): p. 135-143.
4. Liu, M., et al., Glucose starvation triggers filamentous septin assemblies in an S. pombe septin-2 deletion mutant. Biol Open, 2019. 8(1).
5. Gold, V.A., et al., Visualization of cytosolic ribosomes on the surface of mitochondria by electron cryo-tomography. EMBO Rep, 2017. 18(10): p. 1786-1800.
6. Williams, C.C., C.H. Jan, and J.S. Weissman, Targeting and plasticity of mitochondrial proteins revealed by proximity-specific ribosome profiling. Science, 2014. 346(6210): p. 748-51.
doi: https://doi.org/10.1242/prelights.35991
Read preprint (1 votes)
(1 votes) Sign up to customise the site to your preferences and to receive alerts
Register hereAlso in the molecular biology category:
BAF complexes maintain accessibility at stimulus-responsive chromatin and are required for transcriptional stimulus responses
Dina Kabbara
Aging increases ovarian cancer growth, metastasis, and immunosuppression that can be alleviated by inhibiting hedgehog signaling
Zoha Sadaqat
A pre-rRNA positive feedback loop drives malignant ribosome biogenesis
Vaishali Grewal
preLists in the molecular biology category:
Developmental regulation: molecular and ecological niches
This conference was held at the Station Biologique de Roscoff (France) and brought together researchers exploring how diverse niche environments shape developmental processes across scales. Spanning topics from ecological and metabolic influences to signalling networks, mechanics and gene regulation, the meeting highlighted the interplay between intrinsic and extrinsic factors in controlling cell fate and tissue organisation. This preList gathers preprints discussed by speakers and poster presenters during the meeting. Please do get in touch at preLights@biologists.com if you notice any relevant preprints that we may have missed.
| List by | Ingrid Tsang |
preLighters’ choice – Handpicked DevBio preprints
preLighters with expertise across developmental and stem cell biology have nominated a few developmental biology (and related) preprints they’re excited about and explain in a few paragraph why. Concise preprint highlights, prepared by the preLighter community – a quick way to spot upcoming trends, new methods and fresh ideas.
| List by | Theodora Stougiannou et al. |
BSDB Spring Meeting: Molecules to Morphogenesis
The British Society for Developmental Biology (BSDB) Spring Meeting Molecules to Morphogenesis was held from 23–26 March 2026 at the University of Warwick (UK). This meeting brought together a vibrant community of researchers to discuss how molecular mechanisms are integrated across scales to drive morphogenesis, spanning diverse model systems and approaches. This preList contains preprints by presenters from the talk and poster sessions at the meeting. Please do get in touch at preLights@biologists.com if you notice any relevant preprints that we may have missed.
| List by | Ingrid Tsang |
Keystone Symposium on Stem Cell Models in Embryology 2026
The Keystone Symposium on Stem Cell Models in Embryology, 2026, was organised by Jun Wu (UT Southwestern), Jianping Fu (University of Michigan) and Miki Ebisuya (TU Dresden) and held at Asilomar Conference Grounds in California (US). The meeting discussed recent advances made in establishing stem-cell-based embryo models, what fundamental insights into developmental processes have been gleaned from them, as well as how they are beginning to be applied more widely. This prelist contains preprints by presenters at the talk and poster sessions at the conference, which our Reviews Editor in attendance spotted. Please do reach out to preLights@biologists.com if you notice any that we’ve missed.
| List by | Ingrid Tsang |
SciELO preprints – From 2025 onwards
SciELO has become a cornerstone of open, multilingual scholarly communication across Latin America. Its preprint server, SciELO preprints, is expanding the global reach of preprinted research from the region (for more information, see our interview with Carolina Tanigushi). This preList brings together biological, English language SciELO preprints to help readers discover emerging work from the Global South. By highlighting these preprints in one place, we aim to support visibility, encourage early feedback, and showcase the vibrant research communities contributing to SciELO’s open science ecosystem.
| List by | Carolina Tanigushi |
October in preprints – DevBio & Stem cell biology
Each month, preLighters with expertise across developmental and stem cell biology nominate a few recent developmental and stem cell biology (and related) preprints they’re excited about and explain in a single paragraph why. Short, snappy picks from working scientists — a quick way to spot fresh ideas, bold methods and papers worth reading in full. These preprints can all be found in the October preprint list published on the Node.
| List by | Deevitha Balasubramanian et al. |
October in preprints – Cell biology edition
Different preLighters, with expertise across cell biology, have worked together to create this preprint reading list for researchers with an interest in cell biology. This month, most picks fall under (1) Cell organelles and organisation, followed by (2) Mechanosignaling and mechanotransduction, (3) Cell cycle and division and (4) Cell migration
| List by | Matthew Davies et al. |
September in preprints – Cell biology edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading list. This month, categories include: (1) Cell organelles and organisation, (2) Cell signalling and mechanosensing, (3) Cell metabolism, (4) Cell cycle and division, (5) Cell migration
| List by | Sristilekha Nath et al. |
June in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: (1) Cell organelles and organisation (2) Cell signaling and mechanosensation (3) Genetics/gene expression (4) Biochemistry (5) Cytoskeleton
| List by | Barbora Knotkova et al. |
May in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) Biochemistry/metabolism 2) Cancer cell Biology 3) Cell adhesion, migration and cytoskeleton 4) Cell organelles and organisation 5) Cell signalling and 6) Genetics
| List by | Barbora Knotkova et al. |
Keystone Symposium – Metabolic and Nutritional Control of Development and Cell Fate
This preList contains preprints discussed during the Metabolic and Nutritional Control of Development and Cell Fate Keystone Symposia. This conference was organized by Lydia Finley and Ralph J. DeBerardinis and held in the Wylie Center and Tupper Manor at Endicott College, Beverly, MA, United States from May 7th to 9th 2025. This meeting marked the first in-person gathering of leading researchers exploring how metabolism influences development, including processes like cell fate, tissue patterning, and organ function, through nutrient availability and metabolic regulation. By integrating modern metabolic tools with genetic and epidemiological insights across model organisms, this event highlighted key mechanisms and identified open questions to advance the emerging field of developmental metabolism.
| List by | Virginia Savy, Martin Estermann |
April in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) biochemistry/metabolism 2) cell cycle and division 3) cell organelles and organisation 4) cell signalling and mechanosensing 5) (epi)genetics
| List by | Vibha SINGH et al. |
Biologists @ 100 conference preList
This preList aims to capture all preprints being discussed at the Biologists @100 conference in Liverpool, UK, either as part of the poster sessions or the (flash/short/full-length) talks.
| List by | Reinier Prosee, Jonathan Townson |
February in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) biochemistry and cell metabolism 2) cell organelles and organisation 3) cell signalling, migration and mechanosensing
| List by | Barbora Knotkova et al. |
Community-driven preList – Immunology
In this community-driven preList, a group of preLighters, with expertise in different areas of immunology have worked together to create this preprint reading list.
| List by | Felipe Del Valle Batalla et al. |
January in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) biochemistry/metabolism 2) cell migration 3) cell organelles and organisation 4) cell signalling and mechanosensing 5) genetics/gene expression
| List by | Barbora Knotkova et al. |
2024 Hypothalamus GRC
This 2024 Hypothalamus GRC (Gordon Research Conference) preList offers an overview of cutting-edge research focused on the hypothalamus, a critical brain region involved in regulating homeostasis, behavior, and neuroendocrine functions. The studies included cover a range of topics, including neural circuits, molecular mechanisms, and the role of the hypothalamus in health and disease. This collection highlights some of the latest advances in understanding hypothalamic function, with potential implications for treating disorders such as obesity, stress, and metabolic diseases.
| List by | Nathalie Krauth |
BSCB-Biochemical Society 2024 Cell Migration meeting
This preList features preprints that were discussed and presented during the BSCB-Biochemical Society 2024 Cell Migration meeting in Birmingham, UK in April 2024. Kindly put together by Sara Morais da Silva, Reviews Editor at Journal of Cell Science.
| List by | Reinier Prosee |
‘In preprints’ from Development 2022-2023
A list of the preprints featured in Development's 'In preprints' articles between 2022-2023
| List by | Alex Eve, Katherine Brown |
CSHL 87th Symposium: Stem Cells
Preprints mentioned by speakers at the #CSHLsymp23
| List by | Alex Eve |
9th International Symposium on the Biology of Vertebrate Sex Determination
This preList contains preprints discussed during the 9th International Symposium on the Biology of Vertebrate Sex Determination. This conference was held in Kona, Hawaii from April 17th to 21st 2023.
| List by | Martin Estermann |
Alumni picks – preLights 5th Birthday
This preList contains preprints that were picked and highlighted by preLights Alumni - an initiative that was set up to mark preLights 5th birthday. More entries will follow throughout February and March 2023.
| List by | Sergio Menchero et al. |
CellBio 2022 – An ASCB/EMBO Meeting
This preLists features preprints that were discussed and presented during the CellBio 2022 meeting in Washington, DC in December 2022.
| List by | Nadja Hümpfer et al. |
EMBL Synthetic Morphogenesis: From Gene Circuits to Tissue Architecture (2021)
A list of preprints mentioned at the #EESmorphoG virtual meeting in 2021.
| List by | Alex Eve |
FENS 2020
A collection of preprints presented during the virtual meeting of the Federation of European Neuroscience Societies (FENS) in 2020
| List by | Ana Dorrego-Rivas |
ECFG15 – Fungal biology
Preprints presented at 15th European Conference on Fungal Genetics 17-20 February 2020 Rome
| List by | Hiral Shah |
ASCB EMBO Annual Meeting 2019
A collection of preprints presented at the 2019 ASCB EMBO Meeting in Washington, DC (December 7-11)
| List by | Madhuja Samaddar et al. |
Lung Disease and Regeneration
This preprint list compiles highlights from the field of lung biology.
| List by | Rob Hynds |
MitoList
This list of preprints is focused on work expanding our knowledge on mitochondria in any organism, tissue or cell type, from the normal biology to the pathology.
| List by | Sandra Franco Iborra |