








Morphodynamic Atlas for Drosophila Development
Posted on: 10 July 2022
Preprint posted on 27 May 2022
Changes in gene expression and physical form are intricately linked during development, but how do we begin to measure both on a continuous timescale? Mitchell et al provide a path forward with their Morphodynamic Atlas
Selected by Andrew MontequinCategories: developmental biology
Background
Embryos of the fruit fly Drosophila melanogaster have, for over a century, drawn scientists from disparate fields into the study of developmental biology. In recent decades, physical biologists have made strides in understanding the role of morphogen gradients in patterning the body plan (Grimm et al., 2010) as well as the spatial precision with which gene expression patterns can be established and interpreted in the presence of noise (Petkova et al., 2019). These studies have benefited greatly from rigorously controlled quantification of gene expression, as well as the relatively simple geometry of Drosophila embryos. Because of this geometry, it is common to measure the expression levels of gap genes or the concentration of Bicoid with just a single spatial parameter, measuring only distance from the anterior pole of the embryo.
While this one-dimensional view of the embryo has proven useful for both theorists and experimentalists, its limitations become obvious during later morphological events such as gastrulation, where cell flows in multiple directions give physical form to the different germ layers (Streichan et al., 2018). Recent evidence from later stages in Drosophila development indicate that multi-dimensional cell flow may play important roles in generating tissue-level patterning (Gallagher et al., 2022). In this preprint, Mitchell et. al leverage advances in microscopy to generate in toto views of gene expression patterns and cell flows across developmental time and present an analysis pipeline that allows for quantitative comparisons of these features across embryos. This “Morphodynamic Atlas” and the accompanying tools open new doors for studying the relationships between tissue patterning, cell flows, and gene expression patterns. The authors also show that their technique can be applied to later stages of embryogenesis such as organ formation.
Main Findings
The “Morphodynamic Atlas” provides a data set of gene expression and cell flow patterns aligned across space and time
Physical models of development often employ continuous time and space variables, but variation across embryos in developmental timing and tissue deformation can complicate comparisons between these models and experimental data. Mitchell et. al use light sheet microscopy and computational tissue cartography techniques to map the 3-D volumetric imaging data onto a 2-D parameterization of the embryo’s surface. By live imaging the pair-rule gene runt across multiple embryos, they can use its expression pattern as a landmark for building a consensus timeline of development through germ band extension, retraction, and maturation. They also use a similar technique to align images of embryos based on total tissue deformation. Both fixed and live samples can then be compared to this continuous “morphological timeline” to place each image of an embryo at a precise point in developmental time. (Figure 1)
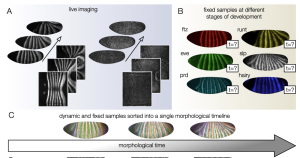
Figure 1: Live images of runt gene expression and a myosin marker (sqhGFP) are spatially and temporally aligned to define a continuous morphological timeline. Fixed images can then be accurately placed along this timeline based on their gene expression and tissue deformation. (Pre-print Fig. 1 A-C)
Drosophila embryonic development is defined by discrete morphological events of stationary cell flow
After generating the atlas of tissue deformation across developmental time, the authors were able to ask how tissue flow patterns evolve during embryogenesis. By comparing instantaneous tissue flow between each imaged time point, the authors observed discrete periods of autocorrelated flow patterns. In other words, the flow patterns across specific ranges of time were highly self-similar. Additionally, these periods of self-similar flow patterns corresponded to previously characterized morphological events such as germ band extension and retraction. These data indicate that tissue flow is quasi-stationary; sources of tissue flow on the embryo’s surface remain fixed during discrete periods of time, with changes in the flow pattern happening in between these morphological events. The authors also found that these flow patterns scale with temperature. Although changes in the temperature can speed up or slow down the overall rate of development, they do not affect the location or direction or tissue flow.
Why I chose this preprint
At first glance, the findings of this paper are not particularly shocking. The “discrete morphological events” of germ band extension and retraction have been well characterized, and temperature variations have long been known to affect the rate of development. Instead, I chose it because of the vision it presents for collecting, analyzing, and interpreting data.
Light sheet microscopy can give unprecedented spatial and temporal resolution of embryonic development, yet the full resolution of the data is rarely ever utilized. The authors of this preprint have developed a toolkit that gives researchers a new language to describe Drosophila development. Rather than reducing development to a handful of stages based on nuclear cycles or qualitative morphological changes, the authors have developed a way to examine changes in shape and gene expression as they naturally occur, on a continuous timescale. In doing so, the authors were able to recapitulate previously understood facets of development, uncover new features of morphological patterning, and provide a rich data set for researchers working to develop field theories of patterning. I am excited to see what discoveries, both experimental and theoretical, are made possible by this technique in the future.
Questions for the authors
- How does distortion from mapping the 3-D volume of the embryo into a 2-D plane affect the alignment? Does distortion at the poles of the embryo mean that small changes in the expression patterns are weighted differently at the poles versus in the middle of the embryo?
- At times when little to no autocorrelation in tissue flow was observed on the whole-embryo scale, are there shorter length scales at which high autocorrelation might be observed? E.g. are there sub-domains within the embryo that might still exhibit stationary flow?
Works Cited
Gallagher, K. D., Mani, M., & Carthew, R. W. (2022). Emergence of a geometric pattern of cell fates from tissue-scale mechanics in the Drosophila eye. ELife, 11. https://doi.org/10.7554/eLife.72806
Grimm, O., Coppey, M., & Wieschaus, E. (2010). Modelling the Bicoid gradient. Development, 137(14), 2253–2264. https://doi.org/10.1242/dev.032409
Petkova, M. D., Tkačik, G., Bialek, W., Wieschaus, E. F., & Gregor, T. (2019). Optimal Decoding of Cellular Identities in a Genetic Network. Cell, 176(4), 844-855.e15. https://doi.org/10.1016/j.cell.2019.01.007
Streichan, S. J., Lefebvre, M. F., Noll, N., Wieschaus, E. F., & Shraiman, B. I. (2018). Global morphogenetic flow is accurately predicted by the spatial distribution of myosin motors. ELife, 7. https://doi.org/10.7554/eLife.27454
doi: https://doi.org/10.1242/prelights.32405
Read preprint (No Ratings Yet)
(No Ratings Yet)Author's response
Sebastian Streichan shared
Thank you for your interest in our work and for contributing this piece. PreLights provides an excellent service to the community!
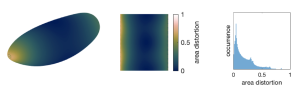
Q1: How does distortion from mapping the 3-D volume of the embryo into a 2-D plane affect the alignment? Does distortion at the poles of the embryo mean that small changes in the expression patterns are weighted differently at the poles versus in the middle of the embryo?
1. The surface of the early embryo is indeed curved, so there is some deformation in the mapping to the plane – just like in a map of the earth. However, because we know the mathematical form of the mapping, we can correct for these deformations whenever we carry out quantitative measurements (e.g. of angles, distances, or regional averages). In contrast with the almost spherical shape of our earth, the fly embryo is nearly cylindrical. This means that curvature – and map deformation – is almost entirely concentrated at the anterior and posterior poles, as shown in the figure below and in Supplementary Figure 2 of Heemskerk & Streichan Nature Methods (2015). Therefore, the clear majority of pixels in our pullbacks are relatively unaffected by map deformation – the surface area corresponding to each pixel varies little, with a standard deviation of <20%. This means that when we work away from the poles, and make measurements only used for time alignment, the map deformations become so weak that they hardly affect the result. We accounted for the variations in dilation across the map in Figure 4 and note that weighting each pixel’s contributions to the r.m.s. velocity changes the resulting curves by only ~3%.
Q2: At times when little to no autocorrelation in tissue flow was observed on the whole-embryo scale, are there shorter length scales at which high autocorrelation might be observed? E.g. are there sub-domains within the embryo that might still exhibit stationary flow?
2. For the 16-hour dataset shown in Figure 3, there is little to no autocorrelation in tissue flow between germ band extension and germ band retraction, and again after retraction. In these
periods, the flow fields at the surface of the embryo are weak, leading to little autocorrelation. However, there are coherent and persistent motions of tissue migration inside the embryo which could potentially exhibit analogous ‘morphodynamic modules’. For instance, after germ band retraction, while the ectodermal motions again slow down, the gut dynamics shown in Figure 5 occur deep inside the embryo and show signs of modular behavior. Later during maturation, there is only noisy autocorrelation at the ectodermal surface. During this time, the embryo experiences rapid muscle contractions, leading to pulsed peristaltic-like motions and rotations which generate noisy signals in the autocorrelation. These would show sustained positive autocorrelation only on much shorter timescales, such as 1-10 seconds. It is intriguing to consider observing development with these fast motions ‘subtracted off’ – i.e. to observe the slow dynamics that persist whilst the embryo’s rapid movement is underway.
3. Finally, we would like to add a comment on where we believe the novelty of our results lies. As you point out, this study builds on a prolific body of work that has produced in-depth analyses of component features (ventral furrow, posterior midgut, cell intercalation/convergent extension) of germ-band extension and retraction. While previous reports have made clear that morphogenesis is stereotypic across individual embryos, to our knowledge the observation that global tissue flow is quasi-stationary for discrete periods during germ-band extension and retraction is novel. Note that identifying a stage by its quasi-stationary flow is altogether different than identifying a stage by the tissue shape; indeed, the shape is continuously changing throughout such a process. We believe the observation of “morphodynamic modules” – enabled by our in-toto approach – raises significant questions about how physical or chemical regulatory inputs can lead to sustained patterns of tissue flow despite mass migration of the underlying cells. Similarly, while it is understood that temperature influences the rate of embryo development, the striking observation that the pattern of tissue deformation only speeds up or slows down as a whole – and remains otherwise undistorted – suggests an astonishing capacity for adaptation, particularly in light of our measurements showing a different scaling for the appearance of mitotic domains. Understanding the molecular bases for these scalings represents exciting future work, as does the question of how different scalings overlay to create reproducible morphogenetic stages across different environmental conditions.
Sign up to customise the site to your preferences and to receive alerts
Register hereAlso in the developmental biology category:
Cell-intrinsic compliance mechanism enables release of tensile stress to prevent tissue rupture
Ruoheng Li
Cooperation between cortical and cytoplasmic forces shapes planar 4-cell stage embryos
Corentin Mollier, Shivani Dharmadhikari
A drought stress-induced MYB transcription factor regulates pavement cell shape in leaves of European aspen (Populus tremula)
Jeny Jose
preLists in the developmental biology category:
Keystone Symposium on Stem Cell Models in Embryology 2026
The Keystone Symposium on Stem Cell Models in Embryology, 2026, was organised by Jun Wu (UT Southwestern), Jianping Fu (University of Michigan) and Miki Ebisuya (TU Dresden) and held at Asilomar Conference Grounds in California (US). The meeting discussed recent advances made in establishing stem-cell-based embryo models, what fundamental insights into developmental processes have been gleaned from them, as well as how they are beginning to be applied more widely. This prelist contains preprints by presenters at the talk and poster sessions at the conference, which our Reviews Editor in attendance spotted. Please do reach out to preLights@biologists.com if you notice any that we’ve missed.
| List by | Ingrid Tsang |
November in preprints – DevBio & Stem cell biology
preLighters with expertise across developmental and stem cell biology have nominated a few developmental and stem cell biology (and related) preprints posted in November they’re excited about and explain in a single paragraph why. Concise preprint highlights, prepared by the preLighter community – a quick way to spot upcoming trends, new methods and fresh ideas.
| List by | Aline Grata et al. |
October in preprints – DevBio & Stem cell biology
Each month, preLighters with expertise across developmental and stem cell biology nominate a few recent developmental and stem cell biology (and related) preprints they’re excited about and explain in a single paragraph why. Short, snappy picks from working scientists — a quick way to spot fresh ideas, bold methods and papers worth reading in full. These preprints can all be found in the October preprint list published on the Node.
| List by | Deevitha Balasubramanian et al. |
October in preprints – Cell biology edition
Different preLighters, with expertise across cell biology, have worked together to create this preprint reading list for researchers with an interest in cell biology. This month, most picks fall under (1) Cell organelles and organisation, followed by (2) Mechanosignaling and mechanotransduction, (3) Cell cycle and division and (4) Cell migration
| List by | Matthew Davies et al. |
June in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: (1) Cell organelles and organisation (2) Cell signaling and mechanosensation (3) Genetics/gene expression (4) Biochemistry (5) Cytoskeleton
| List by | Barbora Knotkova et al. |
Keystone Symposium – Metabolic and Nutritional Control of Development and Cell Fate
This preList contains preprints discussed during the Metabolic and Nutritional Control of Development and Cell Fate Keystone Symposia. This conference was organized by Lydia Finley and Ralph J. DeBerardinis and held in the Wylie Center and Tupper Manor at Endicott College, Beverly, MA, United States from May 7th to 9th 2025. This meeting marked the first in-person gathering of leading researchers exploring how metabolism influences development, including processes like cell fate, tissue patterning, and organ function, through nutrient availability and metabolic regulation. By integrating modern metabolic tools with genetic and epidemiological insights across model organisms, this event highlighted key mechanisms and identified open questions to advance the emerging field of developmental metabolism.
| List by | Virginia Savy, Martin Estermann |
Biologists @ 100 conference preList
This preList aims to capture all preprints being discussed at the Biologists @100 conference in Liverpool, UK, either as part of the poster sessions or the (flash/short/full-length) talks.
| List by | Reinier Prosee, Jonathan Townson |
BSDB/GenSoc Spring Meeting 2024
A list of preprints highlighted at the British Society for Developmental Biology and Genetics Society joint Spring meeting 2024 at Warwick, UK.
| List by | Joyce Yu, Katherine Brown |
GfE/ DSDB meeting 2024
This preList highlights the preprints discussed at the 2024 joint German and Dutch developmental biology societies meeting that took place in March 2024 in Osnabrück, Germany.
| List by | Joyce Yu |
‘In preprints’ from Development 2022-2023
A list of the preprints featured in Development's 'In preprints' articles between 2022-2023
| List by | Alex Eve, Katherine Brown |
preLights peer support – preprints of interest
This is a preprint repository to organise the preprints and preLights covered through the 'preLights peer support' initiative.
| List by | preLights peer support |
The Society for Developmental Biology 82nd Annual Meeting
This preList is made up of the preprints discussed during the Society for Developmental Biology 82nd Annual Meeting that took place in Chicago in July 2023.
| List by | Joyce Yu, Katherine Brown |
CSHL 87th Symposium: Stem Cells
Preprints mentioned by speakers at the #CSHLsymp23
| List by | Alex Eve |
Journal of Cell Science meeting ‘Imaging Cell Dynamics’
This preList highlights the preprints discussed at the JCS meeting 'Imaging Cell Dynamics'. The meeting was held from 14 - 17 May 2023 in Lisbon, Portugal and was organised by Erika Holzbaur, Jennifer Lippincott-Schwartz, Rob Parton and Michael Way.
| List by | Helen Zenner |
9th International Symposium on the Biology of Vertebrate Sex Determination
This preList contains preprints discussed during the 9th International Symposium on the Biology of Vertebrate Sex Determination. This conference was held in Kona, Hawaii from April 17th to 21st 2023.
| List by | Martin Estermann |
Alumni picks – preLights 5th Birthday
This preList contains preprints that were picked and highlighted by preLights Alumni - an initiative that was set up to mark preLights 5th birthday. More entries will follow throughout February and March 2023.
| List by | Sergio Menchero et al. |
CellBio 2022 – An ASCB/EMBO Meeting
This preLists features preprints that were discussed and presented during the CellBio 2022 meeting in Washington, DC in December 2022.
| List by | Nadja Hümpfer et al. |
2nd Conference of the Visegrád Group Society for Developmental Biology
Preprints from the 2nd Conference of the Visegrád Group Society for Developmental Biology (2-5 September, 2021, Szeged, Hungary)
| List by | Nándor Lipták |
Fibroblasts
The advances in fibroblast biology preList explores the recent discoveries and preprints of the fibroblast world. Get ready to immerse yourself with this list created for fibroblasts aficionados and lovers, and beyond. Here, my goal is to include preprints of fibroblast biology, heterogeneity, fate, extracellular matrix, behavior, topography, single-cell atlases, spatial transcriptomics, and their matrix!
| List by | Osvaldo Contreras |
EMBL Synthetic Morphogenesis: From Gene Circuits to Tissue Architecture (2021)
A list of preprints mentioned at the #EESmorphoG virtual meeting in 2021.
| List by | Alex Eve |
EMBL Conference: From functional genomics to systems biology
Preprints presented at the virtual EMBL conference "from functional genomics and systems biology", 16-19 November 2020
| List by | Jesus Victorino |
Single Cell Biology 2020
A list of preprints mentioned at the Wellcome Genome Campus Single Cell Biology 2020 meeting.
| List by | Alex Eve |
Society for Developmental Biology 79th Annual Meeting
Preprints at SDB 2020
| List by | Irepan Salvador-Martinez, Martin Estermann |
FENS 2020
A collection of preprints presented during the virtual meeting of the Federation of European Neuroscience Societies (FENS) in 2020
| List by | Ana Dorrego-Rivas |
Planar Cell Polarity – PCP
This preList contains preprints about the latest findings on Planar Cell Polarity (PCP) in various model organisms at the molecular, cellular and tissue levels.
| List by | Ana Dorrego-Rivas |
Cell Polarity
Recent research from the field of cell polarity is summarized in this list of preprints. It comprises of studies focusing on various forms of cell polarity ranging from epithelial polarity, planar cell polarity to front-to-rear polarity.
| List by | Yamini Ravichandran |
TAGC 2020
Preprints recently presented at the virtual Allied Genetics Conference, April 22-26, 2020. #TAGC20
| List by | Maiko Kitaoka et al. |
3D Gastruloids
A curated list of preprints related to Gastruloids (in vitro models of early development obtained by 3D aggregation of embryonic cells). Updated until July 2021.
| List by | Paul Gerald L. Sanchez and Stefano Vianello |
ASCB EMBO Annual Meeting 2019
A collection of preprints presented at the 2019 ASCB EMBO Meeting in Washington, DC (December 7-11)
| List by | Madhuja Samaddar et al. |
EDBC Alicante 2019
Preprints presented at the European Developmental Biology Congress (EDBC) in Alicante, October 23-26 2019.
| List by | Sergio Menchero et al. |
EMBL Seeing is Believing – Imaging the Molecular Processes of Life
Preprints discussed at the 2019 edition of Seeing is Believing, at EMBL Heidelberg from the 9th-12th October 2019
| List by | Dey Lab |
SDB 78th Annual Meeting 2019
A curation of the preprints presented at the SDB meeting in Boston, July 26-30 2019. The preList will be updated throughout the duration of the meeting.
| List by | Alex Eve |
Lung Disease and Regeneration
This preprint list compiles highlights from the field of lung biology.
| List by | Rob Hynds |
Young Embryologist Network Conference 2019
Preprints presented at the Young Embryologist Network 2019 conference, 13 May, The Francis Crick Institute, London
| List by | Alex Eve |
Pattern formation during development
The aim of this preList is to integrate results about the mechanisms that govern patterning during development, from genes implicated in the processes to theoritical models of pattern formation in nature.
| List by | Alexa Sadier |
BSCB/BSDB Annual Meeting 2019
Preprints presented at the BSCB/BSDB Annual Meeting 2019
| List by | Dey Lab |
Zebrafish immunology
A compilation of cutting-edge research that uses the zebrafish as a model system to elucidate novel immunological mechanisms in health and disease.
| List by | Shikha Nayar |