








A modification to the life cycle of the parasite Trypanosoma brucei
Posted on: 13 August 2019
Preprint posted on 29 July 2019
Article now published in eLife at http://dx.doi.org/10.7554/eLife.66028
A parasite voyage re-visited: Trypanosoma brucei transmission to the tsetse fly is successful upon infection with slender forms of the parasite.
Selected by Mariana De NizCategories: cell biology, epidemiology, microbiology
Background
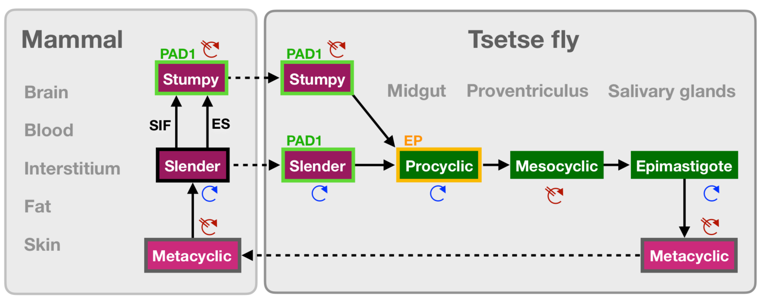
African trypanosomes are the causative parasites of sleeping sickness in humans and nagana in cattle. They are transmitted by tsetse flies upon biting mammalian hosts, given their hematophagous behaviour. In the mammalian host’s circulation, tissues, and interstitium, it is conventionally established that at least two main life cycle stages of the parasite Trypanosoma brucei exist. These stages are known as slender and stumpy, which receive their name from their characteristic morphology. Slender forms undergo a transition into stumpy forms in response to quorum-sensing factors, or stumpy-inducing factors (SIF). This transition is believed to play two important roles: a) to regulate parasite load in the host, preventing hyperparasitemia and early death of the host which would diminish the chance of transmission; and b) to generate a stage that is pre-adapted for survival in the tsetse fly midgut following transmission. Until recently, current knowledge suggested that only stumpy forms were able to infect tsetse flies. The present work by Schuster and Subota et al (1) challenges this paradigm with novel findings that have important epidemiological implications for our understanding of T. brucei transmission from mammalian hosts to tsetse flies.
Key findings and developments
Overall key findings
- Although conventionally thought that only stumpy forms are able to infect tsetse flies, Schuster and Subota et al propose a revision to this concept by showing that slender forms are equally capable of infecting flies.
- While an unanswered epidemiological question is how transmission to tsetse flies can exist when brucei are present in very low parasite densities in mammalian hosts, Schuster and Subota et al show that a single parasite (slender or stumpy) is sufficient to produce an effective infection in the fly.
- Once in the fly, slender parasites can activate expression of the stumpy marker protein associated with differentiation 1 (PAD1) and directly differentiate into procyclic (insect form) stages, without transitioning to the stumpy form and undergoing cell-cycle arrest.
Key tools
- Expression of PAD1 is a proxy for transition into brucei stumpy stages. For most of their work, the authors used a parasite line whereby parasites have GFP positive nuclei when the PAD1 gene is active. This line was used to distinguish slender cells (GFP negative) from stumpy cells (GFP-positive). This was used in combination with nuclear dyes to infer the cell cycle stage of parasites during replication as T. brucei replication follows a sequential process that can be visualized through the parasite nucleus (N) and the kinetoplast (K). In this sequence, T. brucei parasites possess 1K, 1N, followed by 2K, 1N, and finally 2 K, 2N. Although slender parasites are present in all 3 ratios, stumpy forms only exist as 1K1N (thought of as a cell cycle arrest).
- A reporter parasite line was used to study the sequence of events occurring in the fly following ingestion of either stumpy or slender parasites. This line, as the above, encodes the GFP-PAD1 marker, but also an EP1-YFP fusion which allows identifying parasites that begin expressing procyclin on their cell surface, and differentiate to the procyclic life stage.
Specific findings
- The work quantitatively compared completion of the brucei developmental cycle in the tsetse fly, by inducing stumpy production in different ways, namely by a) ectopic expression of a second variant surface glycoprotein (VSG) isoform (i.e. using expression site-attenuated stumpy parasites), and b) parasites produced by incubation with the quorum-sensing stumpy-inducing factor (SIF). Analysis of parasite presence in tsetse flies showed that upon uptake of even a single stumpy form generated by either of the above conditions, led to robust infections in the fly midgut, proventriculus, and salivary glands, in up to 5% of flies.
- Upon feeding flies with PAD1-negative slender trypanosomes from the aforementioned lines, infection efficiency was equal to that observed by stumpy parasite infections.
- Infections with 1-2 slender cells had a higher transmission index than infection with equal numbers of stumpy cells, suggesting that slender cells have at least comparable developmental capacities as the stumpy cells. This finding altogether proposes a revision to the traditional view of the brucei life cycle.
- Infection with slender trypanosomes also begins procyclin expression and differentiates to procyclic forms. Within the fly, by 48-50 h post-infection with slender trypanosomes, the entire population was GFP-PAD1 positive, and a fifth of the population was EP1:YFP positive.
- The authors observed increasing numbers of GFP-PAD1 slender cells, and that these were capable of replicating. This shows that slender parasites activate the PAD1 pathway without being pushed to cell cycle arrest.
- In vitro differentiation experiments using cis-aconitate and temperature drop supported the in vivo observations pointed above.
What I like about this paper
I find this paper very interesting and relevant for parasitology. On one hand because it’s a good example of how unexpected findings can be made based on careful observation and experimentation: it’s important to question established ideas in research fields, and to undertake science that proves or rejects those observations. On the other hand, the finding itself suggests a paradigm-shift for an important piece of knowledge on T. brucei transmission from the mammalian host to the insect vector. This piece of knowledge opens a new research window and could have important implications for our understanding of T. brucei interactions with the vector host, and with the mammalian host at the point of transmission. Equally, while in vitro work is key for research, and has been telling in many parasitology fields, we have often found in various parasites that the relevance and behaviour in vivo, might differ. I find it important that the experiments shown here were performed in relevant vectors.
Open questions
- You found that slender parasites can travel more efficiently than stumpy forms to the salivary glands of the fly. Does infection with stumpy or slender forms have a different effect on tsetse fly fitness/immune response to infection?
- You found that slender forms have a seamless differentiation into procyclic forms without the need to undergo differentiation into stumpy forms. Do procyclic forms arising from slender and from stumpy forms, differ genetically and molecularly?
- While your in vivo experiments in flies open a new research window regarding host-to-vector transmission, what implications does this finding have on how we perceive dynamics, differentiation, and reservoir establishment in the mammalian host? If slender forms and stumpy forms can transmit with equal success and establish an infection in the fly, what is the role of a tissue reservoir- like the skin- in the parasite’s life cycle?
- In a natural feed on an infected host, it is perhaps likely that both stumpy and slender forms are ingested by the fly. Based on your work, slender forms will most successfully invade the salivary glands, while stumpy forms are more able to establish midgut infections (yet have a more limited lifetime). Would ingestion of mixed parasite forms by the fly provide a transmission advantage?
- In your paper, you show that SIF-induced stumpy forms are the fastest in generating procyclic forms, as well as the ones generating procyclics in largest proportions. How do SIF-induced stumpy forms differ from stumpy forms produced in other ways?
- You show that even a single parasite (slender or stumpy) is able to establish infection in the fly. From research in the Plasmodium transmission field, it is known that not every mature gametocyte (the final transmission form) is infectious (2). Moreover, your group has characterized in the fly, various intermediate stages of the parasite’s life cycle in the insect vector (3), while the general concept of the bloodstream forms is that two forms exist: slender forms and stumpy forms. Are intermediate forms transmissible? Are all slender or stumpy forms equally capable of infecting the fly? If not, what is required by brucei to be infectious?
- Following up from the question above, you mention in you discussion that congolense infects tsetse flies without manifesting a cell cycle-arrested stumpy life cycle stage. Are all slender forms of T. congolense equally infectious, and equally capable of establishing infections in the tsetse fly?
- You conclude your pre-print by mentioning that your finding raises the question of what the true biological function of the stumpy life cycle stage actually is. Findings in various parasitology areas show that the production of parasite forms with arrested cycles (like stumpy forms in brucei, or gametocytes in Plasmodium) regulates parasitemia and ensures the survival of the host – as parasite lines which cannot produce such forms kill the host early in infection. In the case of Plasmodium, gametocytes are also stages which are less efficiently recognized by the immune system, establish reservoirs in tissues with less accessibility to drugs (i.e. the bone marrow and the spleen), and are more resistant to many anti-malarial drugs. This capacity of gametocytes ensures Plasmodium transmission despite a) lack of symptoms in the host, and b) clearance of asexual forms of the parasite due to treatment with anti-malarial drugs. Equally, the gametocyte reservoir in the bone marrow ensures a continuous release of mature gametocytes even after parasites in the bloodstream have been cleared. What is known of T. brucei regarding all these aspects, that would make stumpy forms a successful evolutionary adaptation?
References
- Schuster S, Subota I, Lisack J, Zimmermann H, Reuter C, Morriswood B, Engstler M, A modification to the life cycle of the parasite Trypanosoma brucei, bioRxiv, (2019), doi: 10.1101/717975
- Aingaran M, Zhang R, Law SK, Peng Z, Undisz A, Meyer E, Diez-Silva M, Burke TA, Spielmann T, Lim CT, Suresh S, Dao M, Marti M, Host cell deformability is linked to transmission in the human malaria parasite Plasmodium falciparum, Cell Microbiol, (2012), doi: 10.1111/j.1462-5822.2012.01786.x
- Schuster S, Krüger T, Subota I, Thusek S, Rotureau B, Beilhack A, Engstler M, Developmental adaptations of trypanosome motility to tsetse fly host environments unravel a multifaceted in vivo microswimmer system, Elife, (2017) doi: 10.7554/eLife.27656
doi: https://doi.org/10.1242/prelights.13231
Read preprint (No Ratings Yet)
(No Ratings Yet)Sign up to customise the site to your preferences and to receive alerts
Register hereAlso in the cell biology category:
Combinatorial and Inducible CRISPRa/i Enables Canalized hiPSC Forward Programming and Iterative Refinement via Single-Cell Genomics
Cell-ID
Developmental conversion of the nucleolus into an RNA Polymerase II transcriptional platform in Drosophila spermatocytes
Panagiotis Giannios
Cell position is more important than cell shape or age for the acquisition of cell identity in the brown alga Ectocarpus
Urvashi Goswami
Also in the epidemiology category:
Schistosoma haematobium DNA and Eggs in the Urine Sample of School-Age Children (SAC) in South-West Nigeria
Hala Taha
Malaria shaped human spatial organisation for the last 74 thousand years
Alejandra Herbert Leffler's Lab
Green synthesized silver nanoparticles from Moringa: Potential for preventative treatment of SARS-CoV-2 contaminated water
Safieh Shah, Benjamin Dominik Maier
Also in the microbiology category:
Circadian Clock Programming of Anticipatory Antiviral Immunity Gates Enteric Virus Infection Susceptibility
Owen Ang
Inhibition of VP2-mediated entry: a potential antiviral strategy to treat or prevent calicivirus disease
Orestis Savva
Gut microbiome changes over the course of multiple sclerosis differentially influence autoimmune neuroinflammation
Carole Djagang et al.
preLists in the cell biology category:
Developmental regulation: molecular and ecological niches
This conference was held at the Station Biologique de Roscoff (France) and brought together researchers exploring how diverse niche environments shape developmental processes across scales. Spanning topics from ecological and metabolic influences to signalling networks, mechanics and gene regulation, the meeting highlighted the interplay between intrinsic and extrinsic factors in controlling cell fate and tissue organisation. This preList gathers preprints discussed by speakers and poster presenters during the meeting. Please do get in touch at preLights@biologists.com if you notice any relevant preprints that we may have missed.
| List by | Ingrid Tsang |
preLighters’ choice – Handpicked DevBio preprints
preLighters with expertise across developmental and stem cell biology have nominated a few developmental biology (and related) preprints they’re excited about and explain in a few paragraph why. Concise preprint highlights, prepared by the preLighter community – a quick way to spot upcoming trends, new methods and fresh ideas.
| List by | Theodora Stougiannou et al. |
BSDB Spring Meeting: Molecules to Morphogenesis
The British Society for Developmental Biology (BSDB) Spring Meeting Molecules to Morphogenesis was held from 23–26 March 2026 at the University of Warwick (UK). This meeting brought together a vibrant community of researchers to discuss how molecular mechanisms are integrated across scales to drive morphogenesis, spanning diverse model systems and approaches. This preList contains preprints by presenters from the talk and poster sessions at the meeting. Please do get in touch at preLights@biologists.com if you notice any relevant preprints that we may have missed.
| List by | Ingrid Tsang |
Keystone Symposium on Stem Cell Models in Embryology 2026
The Keystone Symposium on Stem Cell Models in Embryology, 2026, was organised by Jun Wu (UT Southwestern), Jianping Fu (University of Michigan) and Miki Ebisuya (TU Dresden) and held at Asilomar Conference Grounds in California (US). The meeting discussed recent advances made in establishing stem-cell-based embryo models, what fundamental insights into developmental processes have been gleaned from them, as well as how they are beginning to be applied more widely. This prelist contains preprints by presenters at the talk and poster sessions at the conference, which our Reviews Editor in attendance spotted. Please do reach out to preLights@biologists.com if you notice any that we’ve missed.
| List by | Ingrid Tsang |
SciELO preprints – From 2025 onwards
SciELO has become a cornerstone of open, multilingual scholarly communication across Latin America. Its preprint server, SciELO preprints, is expanding the global reach of preprinted research from the region (for more information, see our interview with Carolina Tanigushi). This preList brings together biological, English language SciELO preprints to help readers discover emerging work from the Global South. By highlighting these preprints in one place, we aim to support visibility, encourage early feedback, and showcase the vibrant research communities contributing to SciELO’s open science ecosystem.
| List by | Carolina Tanigushi |
November in preprints – DevBio & Stem cell biology
preLighters with expertise across developmental and stem cell biology have nominated a few developmental and stem cell biology (and related) preprints posted in November they’re excited about and explain in a single paragraph why. Concise preprint highlights, prepared by the preLighter community – a quick way to spot upcoming trends, new methods and fresh ideas.
| List by | Aline Grata et al. |
October in preprints – DevBio & Stem cell biology
Each month, preLighters with expertise across developmental and stem cell biology nominate a few recent developmental and stem cell biology (and related) preprints they’re excited about and explain in a single paragraph why. Short, snappy picks from working scientists — a quick way to spot fresh ideas, bold methods and papers worth reading in full. These preprints can all be found in the October preprint list published on the Node.
| List by | Deevitha Balasubramanian et al. |
October in preprints – Cell biology edition
Different preLighters, with expertise across cell biology, have worked together to create this preprint reading list for researchers with an interest in cell biology. This month, most picks fall under (1) Cell organelles and organisation, followed by (2) Mechanosignaling and mechanotransduction, (3) Cell cycle and division and (4) Cell migration
| List by | Matthew Davies et al. |
September in preprints – Cell biology edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading list. This month, categories include: (1) Cell organelles and organisation, (2) Cell signalling and mechanosensing, (3) Cell metabolism, (4) Cell cycle and division, (5) Cell migration
| List by | Sristilekha Nath et al. |
July in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: (1) Cell Signalling and Mechanosensing (2) Cell Cycle and Division (3) Cell Migration and Cytoskeleton (4) Cancer Biology (5) Cell Organelles and Organisation
| List by | Girish Kale et al. |
June in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: (1) Cell organelles and organisation (2) Cell signaling and mechanosensation (3) Genetics/gene expression (4) Biochemistry (5) Cytoskeleton
| List by | Barbora Knotkova et al. |
May in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) Biochemistry/metabolism 2) Cancer cell Biology 3) Cell adhesion, migration and cytoskeleton 4) Cell organelles and organisation 5) Cell signalling and 6) Genetics
| List by | Barbora Knotkova et al. |
Keystone Symposium – Metabolic and Nutritional Control of Development and Cell Fate
This preList contains preprints discussed during the Metabolic and Nutritional Control of Development and Cell Fate Keystone Symposia. This conference was organized by Lydia Finley and Ralph J. DeBerardinis and held in the Wylie Center and Tupper Manor at Endicott College, Beverly, MA, United States from May 7th to 9th 2025. This meeting marked the first in-person gathering of leading researchers exploring how metabolism influences development, including processes like cell fate, tissue patterning, and organ function, through nutrient availability and metabolic regulation. By integrating modern metabolic tools with genetic and epidemiological insights across model organisms, this event highlighted key mechanisms and identified open questions to advance the emerging field of developmental metabolism.
| List by | Virginia Savy, Martin Estermann |
April in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) biochemistry/metabolism 2) cell cycle and division 3) cell organelles and organisation 4) cell signalling and mechanosensing 5) (epi)genetics
| List by | Vibha SINGH et al. |
March in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) cancer biology 2) cell migration 3) cell organelles and organisation 4) cell signalling and mechanosensing 5) genetics and genomics 6) other
| List by | Girish Kale et al. |
Biologists @ 100 conference preList
This preList aims to capture all preprints being discussed at the Biologists @100 conference in Liverpool, UK, either as part of the poster sessions or the (flash/short/full-length) talks.
| List by | Reinier Prosee, Jonathan Townson |
February in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) biochemistry and cell metabolism 2) cell organelles and organisation 3) cell signalling, migration and mechanosensing
| List by | Barbora Knotkova et al. |
Community-driven preList – Immunology
In this community-driven preList, a group of preLighters, with expertise in different areas of immunology have worked together to create this preprint reading list.
| List by | Felipe Del Valle Batalla et al. |
January in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) biochemistry/metabolism 2) cell migration 3) cell organelles and organisation 4) cell signalling and mechanosensing 5) genetics/gene expression
| List by | Barbora Knotkova et al. |
December in preprints – the CellBio edition
A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. This month, categories include: 1) cell cycle and division 2) cell migration and cytoskeleton 3) cell organelles and organisation 4) cell signalling and mechanosensing 5) genetics/gene expression
| List by | Matthew Davies et al. |
November in preprints – the CellBio edition
This is the first community-driven preList! A group of preLighters, with expertise in different areas of cell biology, have worked together to create this preprint reading lists for researchers with an interest in cell biology. Categories include: 1) cancer cell biology 2) cell cycle and division 3) cell migration and cytoskeleton 4) cell organelles and organisation 5) cell signalling and mechanosensing 6) genetics/gene expression
| List by | Felipe Del Valle Batalla et al. |
BSCB-Biochemical Society 2024 Cell Migration meeting
This preList features preprints that were discussed and presented during the BSCB-Biochemical Society 2024 Cell Migration meeting in Birmingham, UK in April 2024. Kindly put together by Sara Morais da Silva, Reviews Editor at Journal of Cell Science.
| List by | Reinier Prosee |
‘In preprints’ from Development 2022-2023
A list of the preprints featured in Development's 'In preprints' articles between 2022-2023
| List by | Alex Eve, Katherine Brown |
preLights peer support – preprints of interest
This is a preprint repository to organise the preprints and preLights covered through the 'preLights peer support' initiative.
| List by | preLights peer support |
The Society for Developmental Biology 82nd Annual Meeting
This preList is made up of the preprints discussed during the Society for Developmental Biology 82nd Annual Meeting that took place in Chicago in July 2023.
| List by | Joyce Yu, Katherine Brown |
CSHL 87th Symposium: Stem Cells
Preprints mentioned by speakers at the #CSHLsymp23
| List by | Alex Eve |
Journal of Cell Science meeting ‘Imaging Cell Dynamics’
This preList highlights the preprints discussed at the JCS meeting 'Imaging Cell Dynamics'. The meeting was held from 14 - 17 May 2023 in Lisbon, Portugal and was organised by Erika Holzbaur, Jennifer Lippincott-Schwartz, Rob Parton and Michael Way.
| List by | Helen Zenner |
9th International Symposium on the Biology of Vertebrate Sex Determination
This preList contains preprints discussed during the 9th International Symposium on the Biology of Vertebrate Sex Determination. This conference was held in Kona, Hawaii from April 17th to 21st 2023.
| List by | Martin Estermann |
Alumni picks – preLights 5th Birthday
This preList contains preprints that were picked and highlighted by preLights Alumni - an initiative that was set up to mark preLights 5th birthday. More entries will follow throughout February and March 2023.
| List by | Sergio Menchero et al. |
CellBio 2022 – An ASCB/EMBO Meeting
This preLists features preprints that were discussed and presented during the CellBio 2022 meeting in Washington, DC in December 2022.
| List by | Nadja Hümpfer et al. |
Fibroblasts
The advances in fibroblast biology preList explores the recent discoveries and preprints of the fibroblast world. Get ready to immerse yourself with this list created for fibroblasts aficionados and lovers, and beyond. Here, my goal is to include preprints of fibroblast biology, heterogeneity, fate, extracellular matrix, behavior, topography, single-cell atlases, spatial transcriptomics, and their matrix!
| List by | Osvaldo Contreras |
EMBL Synthetic Morphogenesis: From Gene Circuits to Tissue Architecture (2021)
A list of preprints mentioned at the #EESmorphoG virtual meeting in 2021.
| List by | Alex Eve |
FENS 2020
A collection of preprints presented during the virtual meeting of the Federation of European Neuroscience Societies (FENS) in 2020
| List by | Ana Dorrego-Rivas |
Planar Cell Polarity – PCP
This preList contains preprints about the latest findings on Planar Cell Polarity (PCP) in various model organisms at the molecular, cellular and tissue levels.
| List by | Ana Dorrego-Rivas |
BioMalPar XVI: Biology and Pathology of the Malaria Parasite
[under construction] Preprints presented at the (fully virtual) EMBL BioMalPar XVI, 17-18 May 2020 #emblmalaria
| List by | Dey Lab, Samantha Seah |
1
Cell Polarity
Recent research from the field of cell polarity is summarized in this list of preprints. It comprises of studies focusing on various forms of cell polarity ranging from epithelial polarity, planar cell polarity to front-to-rear polarity.
| List by | Yamini Ravichandran |
TAGC 2020
Preprints recently presented at the virtual Allied Genetics Conference, April 22-26, 2020. #TAGC20
| List by | Maiko Kitaoka et al. |
3D Gastruloids
A curated list of preprints related to Gastruloids (in vitro models of early development obtained by 3D aggregation of embryonic cells). Updated until July 2021.
| List by | Paul Gerald L. Sanchez and Stefano Vianello |
ECFG15 – Fungal biology
Preprints presented at 15th European Conference on Fungal Genetics 17-20 February 2020 Rome
| List by | Hiral Shah |
ASCB EMBO Annual Meeting 2019
A collection of preprints presented at the 2019 ASCB EMBO Meeting in Washington, DC (December 7-11)
| List by | Madhuja Samaddar et al. |
EMBL Seeing is Believing – Imaging the Molecular Processes of Life
Preprints discussed at the 2019 edition of Seeing is Believing, at EMBL Heidelberg from the 9th-12th October 2019
| List by | Dey Lab |
Autophagy
Preprints on autophagy and lysosomal degradation and its role in neurodegeneration and disease. Includes molecular mechanisms, upstream signalling and regulation as well as studies on pharmaceutical interventions to upregulate the process.
| List by | Sandra Malmgren Hill |
Lung Disease and Regeneration
This preprint list compiles highlights from the field of lung biology.
| List by | Rob Hynds |
Cellular metabolism
A curated list of preprints related to cellular metabolism at Biorxiv by Pablo Ranea Robles from the Prelights community. Special interest on lipid metabolism, peroxisomes and mitochondria.
| List by | Pablo Ranea Robles |
BSCB/BSDB Annual Meeting 2019
Preprints presented at the BSCB/BSDB Annual Meeting 2019
| List by | Dey Lab |
MitoList
This list of preprints is focused on work expanding our knowledge on mitochondria in any organism, tissue or cell type, from the normal biology to the pathology.
| List by | Sandra Franco Iborra |
Biophysical Society Annual Meeting 2019
Few of the preprints that were discussed in the recent BPS annual meeting at Baltimore, USA
| List by | Joseph Jose Thottacherry |
ASCB/EMBO Annual Meeting 2018
This list relates to preprints that were discussed at the recent ASCB conference.
| List by | Dey Lab, Amanda Haage |
Also in the epidemiology category:
BioMalPar XVI: Biology and Pathology of the Malaria Parasite
[under construction] Preprints presented at the (fully virtual) EMBL BioMalPar XVI, 17-18 May 2020 #emblmalaria
| List by | Dey Lab, Samantha Seah |
1
COVID-19 / SARS-CoV-2 preprints
List of important preprints dealing with the ongoing coronavirus outbreak. See http://covidpreprints.com for additional resources and timeline, and https://connect.biorxiv.org/relate/content/181 for full list of bioRxiv and medRxiv preprints on this topic
| List by | Dey Lab, Zhang-He Goh |
1
Antimicrobials: Discovery, clinical use, and development of resistance
Preprints that describe the discovery of new antimicrobials and any improvements made regarding their clinical use. Includes preprints that detail the factors affecting antimicrobial selection and the development of antimicrobial resistance.
| List by | Zhang-He Goh |
Also in the microbiology category:
SciELO preprints – From 2025 onwards
SciELO has become a cornerstone of open, multilingual scholarly communication across Latin America. Its preprint server, SciELO preprints, is expanding the global reach of preprinted research from the region (for more information, see our interview with Carolina Tanigushi). This preList brings together biological, English language SciELO preprints to help readers discover emerging work from the Global South. By highlighting these preprints in one place, we aim to support visibility, encourage early feedback, and showcase the vibrant research communities contributing to SciELO’s open science ecosystem.
| List by | Carolina Tanigushi |
BioMalPar XVI: Biology and Pathology of the Malaria Parasite
[under construction] Preprints presented at the (fully virtual) EMBL BioMalPar XVI, 17-18 May 2020 #emblmalaria
| List by | Dey Lab, Samantha Seah |
1
ECFG15 – Fungal biology
Preprints presented at 15th European Conference on Fungal Genetics 17-20 February 2020 Rome
| List by | Hiral Shah |
EMBL Seeing is Believing – Imaging the Molecular Processes of Life
Preprints discussed at the 2019 edition of Seeing is Believing, at EMBL Heidelberg from the 9th-12th October 2019
| List by | Dey Lab |